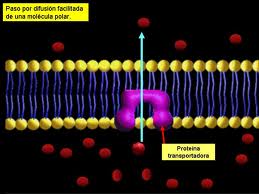
Membrana Plasmática.
· Alberts et al. Biología Molecular de la Célula. 3a. edición, Ed. Omega. 1994.
· Avers, Ch. Biología Celular. 2da. Edición. Grupo Ed. Iberoamérica, S.A. de C.V. 1991.
· Janmey PA, Kinnunen PKJ. Byophisical properties of lipids and dynamic membranes. 2006. Trends in cell biology. 16:538-546.
· Lodish, Berk et al. Biología Celular y Molecular. 4ta edición. Ed. Panamericana. 2002. Madrid España.
· Van Meer G, Voelker DR, Feigenson GW. Membrane lipids: where are they a how they behave. 2008. Nature reviews in molecular cell biology. 9:112-124.
· Singer J. and Nocolson G. The Fluid Mosaic model of the tructure of Cell Membranes. Science. 1972. 175: 720-731.
· Apuntes de la Càtedra Fundamentos de Biologìa. Instituto Superior del Profesorado de Salta. Argentina.
· Castro, Roberto; Handel, María; Rivolta, Graciela: "Actualizaciones en Biologìa" Ed. EUDEBa. 1994.
· http://www.botanica.cnba.uba.ar/Pakete/3er/LaCelula/TransportedeMembrana.htm
· http://www.scribd.com/doc/37501201/Transporte-de-Membrana
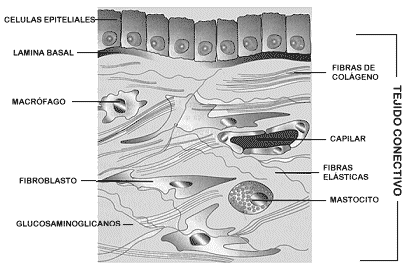
Uniones Celulares y Matriz Extracelular.
· Alberts et al. Biología Molecular de la Célula. 3a. edición, Ed. Omega. 1994.
· Alberts, B. , Bray, D., Roberts, K. y Watson, J. Cell adhesion, cell junctions and the extracellular matrix. In Molecular Biology of Cell, 2ª ed., 1989: 791-836.
· Avers, Ch. Biología Celular. 2da. Edición. Grupo Ed. Iberoamérica, S.A. de C.V. 1991.
· Lodish, Berk et al. Biología Celular y Molecular. 4ta edición. Ed. Panamericana. 2002. Madrid España.
· Van Der Rest, M. Collagen family of protein. FASEB 1991; 5: 2814-2823.